Думата мускул има латински произход - musculus означава мишка в умалителна форма или мишле. Това е основната анатомична единица на мускулната система, част от опорно - двигателния апарат на висшите животни. Образува се от средния зародишен лист на ембрионите. Мускулите са еднакво устроени органи, които имат способността да се съкращават и разпускат. Функцията на мускулите е да упражняват статично въздействие или да предизвикват движение на части от тялото, включително на вътрешните органи. Движенията на тялото са резултат от съвместната работа на нервната и мускулната система и представляват една от най-важните разлики между животните и растенията.
Мускулите са изградени от множество мускулни влакна, които имат способността да се съкращават. Мускулните влакна са клетки със сравнително просто устройство: съдържат само клетъчно ядро и голяма цитоплазма. Енергията за мускулните съкращения се получава от химичното съединение аденозинтрифосфат (съкратено АТФ). Мускулите се делят на скелетни, сърдечни и гладки. Съкращаването на сърдечните и гладките мускули протича неволево и работата им е необходима за оцеляването на организма. Примери за това са работата на сърцето и червата. Волевото съкращаване на скелетните мускули се използва за придвижване. Мускулите се захранват главно чрез окисление на мазнини и въглехидрати, но понякога и анаеробно. В резултат на тези химични реакции се получават молекули аденозинтрифосфат. Всеки път по време на физическо натоварване се образуват микроскопични увреждания (разкъсвания) на мускулните влакна и връзките. След това обаче тялото ги възстановява и те стават още по-здрави, по-силни, по-големи и по-издръжливи. Именно на този принцип, чрез редовни физически тренировки мускулите нарастват, а сухожилията се заздравяват.
Мускулната тъкан е образувана от силно удължени клетки, наречени мускулни влакна. Миофибрилата е основната функционална единица и е изградена от съкратителни белтъци – актин и миозин. Основната функция на мускулната тъкан е двигателната, която осигурява възбудимостта, проводимостта и съкратимостта. Възбудимост е свойството на клетката да премине от състояние на покой в активно състояние под действие на дразнители. Проводимост е способността на клетката да предава възбуждането като нервен импулс по дължината си. Съкратимост е способността на мускулната клетка да намалява дължината си. Намаляването на дължината на всеки един мускул е единственият начин да се генерира сила от живата материя - не съществува мускул, който да може да се разтяга волево. Екстензията на мускулите е чисто пасивна - чрез свиване на мускулите - антагонисти, които разтягат ставите и костите, при което противодействащият мускул също увеличава своята дължина.
Мускулните клетки подобно на невроните могат да бъдат възбудени от различни химични, механични и електрически дразнители, при което се генерира акционен потенциал. Този потенциал се провежда по хода на клетъчната мембрана. За разлика от неврона обаче мускулната клетка има съкратителни свойства, които се проявяват под действие на акционния потенциал. Всяко мускулно влакно е отделна клетка - удължена, цилиндрична, многоядрена, обвита от клетъчна мембрана, наречена сарколема. Между съседните клетки няма синцитиални мостчета. Мускулните клетки съдържат фибрили, които са съставени от отделни филаменти. Последните са изградени от съкратителни белтъци. Съкращението на мускула зависи основно от следните белтъчни молекули:
миозин - молекулно тегло 460 000
актин - молекулно тегло 43 000
тропомиозин - молекулно тегло 70 000
тропонин - той е изграден от три субединици - тропонин I, T и C. Те имат молекулни тегла между 18 000 и 35 000
алфа - актинин - молекулно тегло 190 000. Той прикрепва актина към Z-линиите
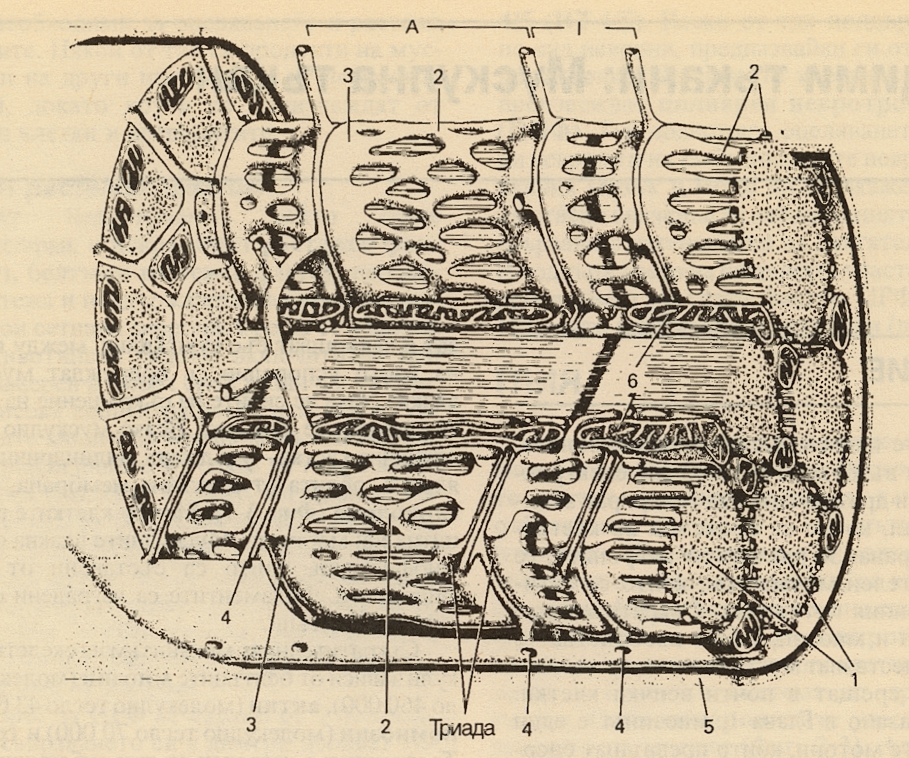
Произходът на мускулната тъкан е мезодермален. Скелетната мускулна тъкан е образувана от влакна, които под микроскоп изглеждат напречнонабраздени. Изгражда мускулите на туловището, крайниците и главата. Чрез съкращението ѝ се осъществява придвижването на тялото в пространството. Съкращенията на напречнонабраздената мускулна тъкан са волеви – настъпват по желание на човека. Мускулатурата на скелета е изградена от самите мускулни влакна и малко количество рехава влакнеста тъкан, която огражда мускулните съдове и нерви. Отвън мускулите са покрити от дебела капсула от уплътнена влакнеста съединителна тъкан - епимизиум. Напречнонабраздената мускулна тъкан има дълги многоядрени клетки, като ядрата са разположени в периферията на клетките, под клетъчната мембрана. При наблюдение с микроскоп на напречнонабраздената мускулна тъкан клетките имат тъмни и светли ивици, което се дължи на миофибрилите - от там тъканта е получила и името си. Тези мускулни влакна изграждат скелетните мускули и извършват бързи и мощни съкращения с кратък период на почивка, затова се изморяват лесно. Съкращават се волево под действието на импулси, идващи от кората на главния мозък. Заемат 40 % от масата на организма, като при силови атлети с по-развит опорно - двигателен апарат този процент се повишава значително. Причината за характерното набраздяване на скелетната мускулатура е разликата в индекса на пречупване на светлината, характерни за различните части на мускулното влакно. Отделните елементи на напречното набраздяване се отбелязват с латински букви. През средата на светлата ивица I преминава тъмната Z-линия, а тъмната ивица А има в средата си по-светла Н-ивица. В средата на тази Н-ивица има напречна М-линия и тази линия заедно с тесните светли области от двете ѝ страни понякога се наричат псевдо Н-зона. Участъкът между двете съседни Z-мембрани се нарича саркомер. На долната схема е представено подреждането на тънките и дебелите филаменти, на което се дължи напречната набразденост. Дебелите филаменти имат около два пъти по-голям диаметър от тънките и са изграднеи от миозин, а тънките са изградени от актин, тропомиозин и тропонин. Миозиновите филаменти образуват споменатите вече А-ивици, докато подреждането на тънките води до образуването на по-малко плътните I-ивици. По-светлите Н-зони в центъра на А-ивиците са области, в които при отпуснат мускул тънките филаменти не се застъпват с дебелите. Z-линиите пресичат фибрилите и са свързани с тънките филаменти. При електронномикроскопско изследване се вижда че около всеки дебел филамент се разполагат 6 тънки филамента, подредени в правилна хексагонална структура.
Схематично представяне на скелетен мускул от бозайник. Единичното мускулно влакно е обвито в сарколема. С цифрата 1 е означен напречният срез, който показва отделните миофибрили. Дебелите филаменти имат около два пъти по-голям диамтър от тънките и са изградени от миозин. Тънките са изградени от актин, тропонин и тропомиозин. Дебелите са подредени така че образуват А-ивиците, докато подреждането на тънките филаменти води до подреждането на по-малко плътните I-ивици. Т - системата и прилежащите ѝ цистерни от саркоплазмения ретикулум образуват триади. Базалната ламина е означена с цифрата 5 и обгръща цялата сарколема. С цифрата 6 са означени митохондриите.

Електронномиркоскопска снимка на човешки m. gastrocnemius. Отделните ивици и линии са означени със сътоветните букви над образа. Миозинът е сложен белтък, който може да се свързва с актина. Формата, която се среща в мускулите, е миозин 2, с две глобуларни глави и дълга опашка. Миозин 2 се среща и в други клетки заедно с миозин 1, който има само една глава. Молекулата му е изградена от две тежки и четири леки вериги, като леките вериги и N-крайните части на тежките вериги се свързват и образуват глобуларни глави. Чисто схематично това изглежда така:
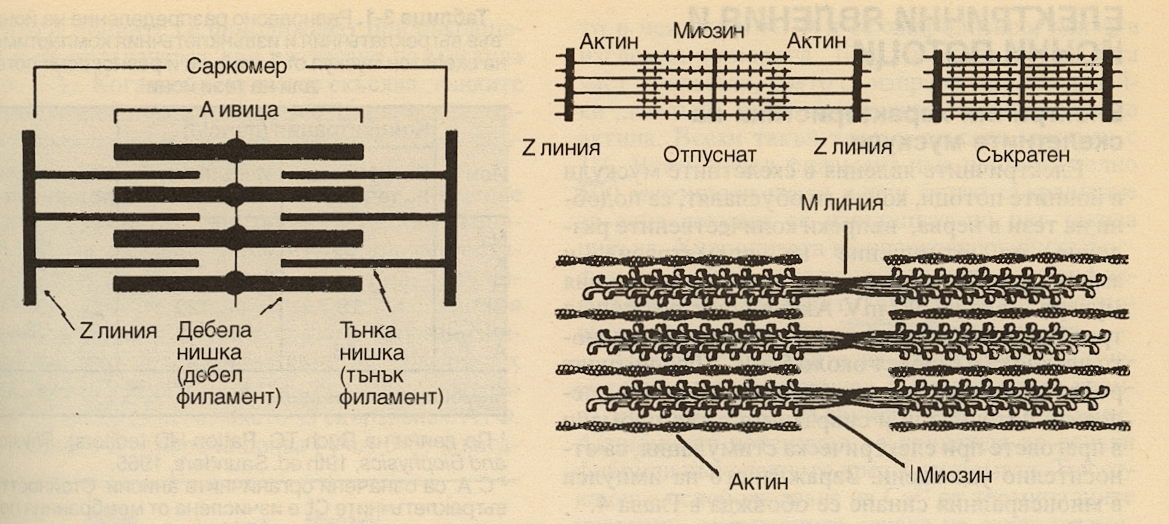
Вляво е показано подреждането на тънки (актинови) и дебели (миозинови) филаменти в скелетен мускул. Вдясно е показано приплъзването на актина спрямо миозина по време на мускулното съкращение и приближаването на Z-линиите. Напречната набразденост на мускулите е възникнала още в ранните етапи на еволюцията. Различните типове скелетно-мускулни влакна могат да се класифицират на базата на максималната си скорост на скъсяване, според което те се делят на бързи и бавни влакна. Двата вида влакна съдържат изоензими, които се различават по начина, по който разцепват АТФ. Това определя скоростта за максималното скъсяване на мускулните влакна. Влакна, съдържащи миозин с висока АТФ-активност са бързи, а тези с миозин с по-ниска АТФ-активност са бавни. Бързите мускулни влакна се съкращават бързо, но за сметка на това се уморяват бързо.
Според вида на главния път за образуване на АТФ влакната се делят на оксидативни и гликолитични. Първият тип съдържат множество митохондрии и в резултат на това имат висок капацитет на окислително фосфорилиране. Заобиколени са от множество кръвоносни съдове и съдържат големи количества от протеина миоглобин. Последният придава тъмночервен цвят на мускулите и осигурява малки кислородни запаси във влакното. Гликолитичните влакна имат малко количество митохондрии, но за сметка на това съдържат голяма концентрация на гликолитичен ензим и имат големи запаси от гликоген. Заобиколени са от по-малко кръвоносни съдове и съдържат по-малко количество миоглобин. Това обезцветява влакното, поради което тази тъкан се нарича бяла мускулатура. Обикновено гликолитичните влакна имат по-голям диаметър от оксидативните. Болшинството мускули са смесени, като в различни пропорции съдържат бели и червени влакна. По-ясно разделение на бели и червени мускули например има при заека и кокошката. При птиците съществуват дорзални мускули, които са изградени почти само от оксидативни мускулни влакна - тъй като е необходима голяма издържливост на мускулатурата по време на полет, в процеса на еволюцията мускулатурата се е оформила именно по такъв начин. При бързите гликолитични влакна миозиновата аденозинтрифосфатаза има висока активност, докато при бавните окислителни влакна тази актовност е на средно ниво.
Разликите между различните мускулни влакна са обусловени от различните белтъци в тях. Описани са около 10 форми на тежките вериги на миозина, като те се отличават по отношение на аминокиселинния си състав. При зрели хора съотношението между отделните изоформи определя характеристиките на всеки един мускул. Освен това тези промени могат да се определят от изменения на активността, инервацията и нивата на различни хормони. Бързите влакна могат да се превръщат в бавни и обратно.
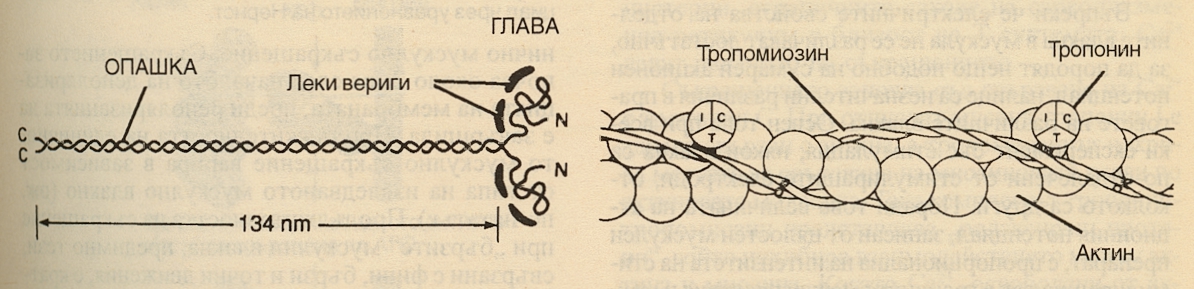
Вляво е показана молекулата на миозин 2, която е изградена от завити помежду си две тежки и четири леки вериги. Вдясно пък е представено подреждането на актина, тропомиозина и трите субединици на тропонина - I, C и T. Глобуларните глави съдържат място, към което се свързва актин и каталитично място са свързване на АТФ. Молекулите са подредени така че главите на миозиновите молекули образуват напречни мостчета към актиновите. Миозиновите молекули са подредени симетрично от двете страни на центъра на саркомера и това подреждане обуславя светлите области в псевдо Х-зоната. М-линията се дължи на централно издуване на всеки от дебелите филаменти. На тези места се намират тънки напречни връзки, които поддържат точното подеждане на дебелите филаменти. Във всеки дебел сегмент има по няколкостотин миозинови молекули. Молекулите на тропонина са малки глобуларни единици, разположени на определени интервали по дължината на тропомиозиновите вериги. Тропонин Т свързва към тропомиозина останалите тропонинови компоненти; тропонин I инхибира взаимодействието на миозина с актина, а тропонин С съдържа свързващите места за калциевите йони и дава началото на мускулното съкращение.
Тънките филаменти са изградени от две вериги от глобуларни единици, формиращи дълга двойна спирала. В браздата между двете актинови вериги е разположена дългата нишковидна молекула на тропомиозина, както е показано на схемата горе. Всеки тънък филамент съдържа 300 - 400 актинови и 40 - 60 тропомиозинови молекули.
Миофибрилите са заобиколени от структури, изградени от мембрани, които на електронномикроскопски снимки изглеждат като мехурчета и тръбички. Тези структури образуват саркотубулната система, която се състои от Т-система и саркоплазмен ретикулум. Т-системата от напречни каналчета е продължение на мембрната на мускулното влакно и образува решетка, перфорирана от отделните миофибрили. Пространството между двата пласта на Т-системата е продължение на екстрацелуларното пространство. Саркоплазменият ретикулум, който образува неравномерната неравномерна обвивка около всяка фибрила, има удължени терминални цистерни, разположени в непосредствена близост с Т-системата между ивиците А и I. В тези места на контакт подреждането на центрлната Т-система с разположените от двете ѝ страни цистерни от саркоплазмения ретикулум е довело до въвеждането на термина триади. Функцията на Т-системата, която е продължение на сарколемата, е провеждането на акционния потенциал от клетъчната мембрана до всички фибрили в мускула. Саркоплазменият ретикулум има значение за движението на калциевите йони и за мускулния метаболизъм.
Гладката мускулатура се състои от къси, гладки влакна, при които липсва напречната набразденост. Изграждат кухите вътрешни органи – стомах, черва, кръвоносни съдове и други. От съкращението им зависят жизненоважни процеси като дишането, кръвообращението и храносмилането. Съкращението на гладката мускулна тъкан е неволево – не се извършва в зависимост от желанието на човека. Контрахира се бавно, но развива значителна сила. Изградена е от вретеновидни клетки с едно ядро, разположено в центъра. Миофибрилите трудно се различават под микроскоп, поради което и гладката мускулна тъкан е наречена по този начин. Извършва бавни и ритмични съкращения без признаци на умора и за разлика от напречнонабраздената се контролира от вегетативната нервна система. Повишаването на температурата води до разширяването на влакната, а студът стимулира съкращението им.
Най-голямата разлика между гладката и напречнонабраздената мускулатура е при контрактилните протеини. При скелетните мускули те са прецизно подредени, в резултат на което се наблюдава и набраздеността. При гладките липсва такава прецизност при подреждането, поради което липсва и набразденост. Гладките мускули се контрахират много по-бавно от скелетните, скоростта на разпространение на възбудата е също по-ниска. Съкращаването им и поддържането на тонуса се извършва с много по-малко енергия в сравнение с напречнонабраздените. Ето защо въпреки че работят непрекъснато през целия живот, гладките мускули не се уморяват.
Сърдечната мускулатура е образувана от влакна, които са по-къси от тези, които образуват скелетната мускулна тъкан. Поради това под микроскоп те също изглеждат напречнонабраздени. Сърдечната мускулна тъкан изгражда мускулите на сърцето. Заедно с гладката мускулна тъкан тя се причислява към неволевата мускулатура. Според различни конспиративни теории някои добре тренирани йоги могат по собствено желание да си спират сърдечната дейност, което от медицинска гледна точка представлява чиста глупост. Въпреки това обаче сърдечната мускулатура притежава известна автономия и автоматизъм. Ритмичните контракции се дължат на собствен команден пункт - синусов възел. Въпреки това обаче важна роля в регулацията на сърдечната дейност изпълнява нервната тъкан. Допълнително сърдечната мускулна тъкан се изгражда от два подвида – работна и проводяща. Изградена е от клетки, които в краищата си се разклоняват и се свързват чрез междуклетъчни дискове. Извършва бързи и ритмични съкращения с дълъг период на почивка, затова и не се изморява. За нея е характерно свойството автоматизъм – способността да генерира импулси без въздействие отвън, но същевременно се влияе и от вегетативната нервна система. Среща се само в областта на сърцето.
Скелетните мускули при бозайниците съдържат вода (72 – 80 %), белтъчини (16.5 – 21 %), мазнини и липиди (3.9 %). Белтъците на мускулатурата се делят на две групи: белтъци на саркоплазмата (миоген, глобулин Х, миоглобин) и белтъци на миофибрилите – миозин, актомиозин, актотропомиозин, тропомиозин. Миоглобинът е белтъкът, който осигурява запасите на мускула с кислород, като се свързва с него. Така се превръща в оксимиоглобин. Продължително работещите мускули съдържат повече миоглобин. Миоглобинът се открива в по-голямо количество при животните, които издържат повече време под водата без да дишат. Така например при тюлена 47 % от кислородните запаси в организма са свързани с миоглобина, докато при други сухоземни животни този процент е от порядъка на 13 – 16 %. Молекулата на миоглобина се състои от една полипептидна верига, съдържаща хем, който е активният център за свързване на кислорода. Миоглобинът има висока афинитет към кислорода, което му позволява да свързва кислорода ефективно в условия на ниско съдържание на кислород, например в мускулите по време на физическа активност. Когато мускулите се нуждаят от кислород, миоглобинът го освобождава от връзката и го предава на митохондриите вътре в мускулите, където се осъществява окислително фосфорилиране. Така се осигурява енергия за мускулните съкращения. Миоглобинът също играе важна роля в диагностиката на сърдечни и мускулни заболявания. Повишени нива на миоглобин в кръвта могат да бъдат индикатор за увреждане или некроза на мускулите - дори за инфаркт на миокарда. Миоглобинът може също да се използва като биомаркер за оценка на травмите на мускулите при физическа активност или след прием на определени медикаменти.
Големината, формата и размера на мускула са твърде разнообразни и зависят от разположението и функцията, която извършват. Скелетните мускули обикновено притежават една централна активна част, наречена мускулно тяло или корем и крайни механични части – сухожилия. Чрез последните се закрепват за скелета. По външен вид мускулите могат да бъдат дълги, къси и плоски, като се обединяват в няколко групи:
Вретеновидни мускули. Имат правилна вретеновидна форма с кръгъл или леко сплеснат разрез. Срещат се предимно в крайниците - например двуглавият мускул на мишницата (бицепс) е типичен пример за това, макар че мускулът може да се причисли и към групата на двуглавите
Многоразделни мускули. Представляват дълга и различно дебела мускулна маса. Чрез многобройни къси сухожилия се залавя за множество кости. Този вид мускули се срещат около гръбначния стълб
Лентовидни мускули. Формата им е на различно широки плоски ленти, които съединяват отдалечени кости. Срещат се по долната повърхност на шията
Плоски мускули. Представляват сравнително тънки мускулни плочи с голяма повърхност и различна форма. Обикновено мускулните снопчета завършват в широко тънко сухожилие. Една част от тези мускули образуват стената на коремната кухина.
Зъбчати мускули. Съставени са от голям брой мускулни лъчи или триъгълни зъбци, които се залавят за последователно наредени костни образувания като прешлени и ребра
Пръстеновидни мускули. Представляват затворен пръстен, който при контракция затваря някои отвори на тялото – устен, анален отвор, клепачи
Двуглави, триглави и четириглави мускули. Започват от две, три или четири глави и накрая завършват в общо крайно сухожилие
Къси мускули. Това са мускули с ограничена дължина и различна форма. Съединяват отделни части на две съседни кости. Намират се около гръбначния стълб и костите на китката и ходилото
Общи мускули. Започват с една или повече сраснали мускулни глави, които преминават в общо сухожилие. Впоследствие то се разделя на толкова сухожилия, колкото пръсти има организмът
Двукоремни мускули. Това са мускули съставени от две вретеновидни тела, разположени надлъжно едно на друго и разделени помежду си от междинно сухожилие.
Според разположението в тялото мускулите се делят на следните няколко групи:
Мускули на главата:
Мимически мускули – прикрепени са от едната си страна за костите на черепа, а за другата са вплетени в дълбокия слой на кожата. При съкращаването си тези мускули променят изказа на лицето. Те са разположени около очите, устата и носа. При движенията си отварят и затварят очните цепки, движат устните, стесняват и разширяват ноздрите
Дъвкателни мускули – те са по 4 от всяка страна на лицето. Започват от черепа и се залавят за долната челюст. При движението си осигуряват дъвкателни движения
Мускули на шията – чрез тях се извършва движението на главата и шията
Мускули на туловището:
Гръбни мускули
Гръдни мускули
Коремни мускули.
Мускули на крайниците:
Мускули на горния крайник
Мускули на долния крайник.
Според действието са описани следните групи мускули:
Агонисти (Agonist muscles) – мускули, извършващи еднакво действие - сгъване, привеждане, завъртане
Антагонисти (Antagonist muscles) – мускули, извършващи противоположно действие, пр. едните сгъват, а другите разгъват
Синергисти (Synergists) – мускули, които участват в движението индиректно. Те могат да участват при стабилизиране на ставните връзки, така че друго движение да бъде извършено (стабилизиращи мускули).
Според доц. д-р Стоян Гаджев единна класификация на скелетните мускули няма и всеки мускул може да бъде класифициран към следните групи:
А. По положение:
1. Повърхностен
2. Дълбок
3. Външен
4. Вътрешен
5. Латерален
6. Медиален
Б. По форма:
1. Вретеновиден мускул
2. Лентовиден мускул
3. Плосък мускул
4. Триъгълен мускул
5. Квадратен мускул
6. Пръстеновиден мускул
7. Зъбчат мускул
8. Трапецовиден мускул
9. Ромбовиден мускул
В. По големина:
1. Голям
2. Малък
3. Дълъг
4. Къс
Г. По отношение на ставите:
1. Едноставен мускул
2. Двуставен мускул
3. Многоставен мускул
Д. Други мускулни форми:
1. Двукоремен мускул
2. Многокоремен мускул, m. digastricus
3. Името на мускула произлиза от неговото начало и край (m. brachiocephalicus)
4. Мускулът започва от една кост, след това се разделя и отделните части завършват с отделни сухожилия на няколко места (пръстовите свивачи и изпъвачи)
Е. Според функцията, която мускулът изпълнява:
1. Свивач
2. Изпъвач
3. Отвеждач
4. Привеждач
5. Завъртач
5.1. Пронатор
5.2. Супинатор
6. Разширител
7. Изопвач (обтегач)
8. Спускач (теглещ надолу)
9. Повдигач
10. Ретрактор
11. Протрактор
12. Стеснител
13. Орбикуларен
14. Ставен
Ж. Мускул, при който влакната се свързват под ъгъл със сухожилието може да бъде:
1. Едноперест мускул
2. Двуперест мускул
3. Многоперест мускул
Същността на мускулното съкращение е сложна поредица от биохимични процеси. Съкращаването и последващото разпускане може да се схематизира по следния начин: повърхността на мускулната мембрана на влакното се деполяризира от акционния потенциал. Сигналът се отвежда дълбоко във влакното посредством Т-каналчестата система. Електрическият сигнал се разпростира от Т-каналчетата към саркоплазматичния ретикулум. Сигналът предизвиква освобождаване на калциеви йони, които се намират в ретикулума. Нараства концентрацията на калциеви йони в саркоплазмата, те се свързват с тропонина и се осъществяват конформационни промени. Предизвиква се промяна на тропомиозиновите молекули. В резултат на това миозиновите мостчета въздействат върху актиновите нишки. След като се допират до актиновите нишки миозиновите мостчета чрез своите движения задвижват и тях. Извършва се плъзгане на актиновите нишки - те навлизат в анизотропните дискове между миозиновите нишки. Така се скъсява саркомерът и съкращаването е завършено. Калциевите йони от саркоплазмата се свързват със саркоплазмения ретикулум. Тропомиозинът инхибира допирането на мостчетата до актиновите нишки и мускулът се отпуска до следващата деполяризация. Потенциалът на покой на скелетния мускул е около -90 mV. Акционният потенциал продължава 2 - 4 милисекунди и се провежда по мускулното влакно със скорост около 5 метра в секунда. Абсолютният ефрактерен период продължава 1 - 3 милисекунди, а следовите потенциали и свързаните с тях промени в праговете при електрическата деполяризация са относително удължени.
Разпределението на йоните от двете страни на мембраната на мускулната клетка е подобно на това при нервната. Натриевите катиони са в много по-голямо количество извън клетката, докато при калиевите е обратното. Както и при нервите, деполяризацията е в резултат на навлизане на натрий, а реполяризацията - излизане на калий. Важно е да се отличат електричните от механичните явления в мускула - въпреки че едните не могат да съществуват без другите, техните физиологични характеристики са различни. Деполяризацията на мускулното влакно започва от моторната плочка - специализирана структура, която се намира под окончанието на двигателния нерв. От там деполяризацията се разпространява по хода на цялото влакно и дава началото на съкратителния отговор. Единичен акционен потенциал предизвиква бързо съкращение, последвано от бързо отпускане. Графично този процес изглежда така:
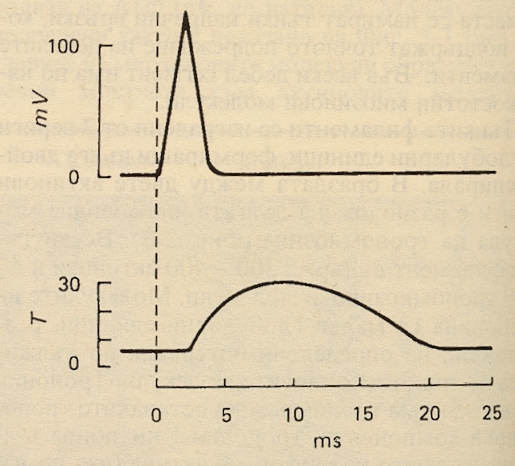
На горната графика е показан електричният отговор, а на долната - механичният при генериране на акционен потенциал в мускул на бозайник. Абсцисната ос е една и съща - на нея е нанесено времето в милисекунди. Мускулната клетка притежава свойството дразнимост, като специфичният отговор на мускулите при дразнене е тяхното съкращение. То се предизвиква по два начина – пряко, като се действа с дразнител направо върху мускула и нормално, когато мускула се възбужда след дразнене на съответния нерв, който осигурява двигателната инервация. Мускулните дразнения могат да бъдат естествени (присъщи) и изкуствени (неприсъщи). Изкуствените биват механични, физични и химични. Естествените дразнения за мускула са импулсите, които се предават върху него посредством съответния нерв. Мускулната възбудимост се променя под влияние на различни фактори. Подобрява се чрез подходящи упражнения, които активират метаболитните процеси. Възбудимостта се понижава след продължителна почивка или много активна мускулна дейност, водеща до умора. За мускулната тъкан са характерни фазовите явления на възбудния процес, които са с различна продължителност. Наблюдава се рефрактерна фаза, екзалтационна фаза, фаза на понижена възбудимост и накрая мускулът изпада в състояние на нормална възбудимост. В зависимост от функцията си мускулите се делят на изпъвачи, свивачи, отдалечаващи, приближаващи, ротиращи и други. Тези различни мускули на тялото, които изпълняват една и съща функция или се подпомагат взаимно се наричат синергисти, а когато изпълняват противоположни функции се наричат антагонисти. Латентен период се нарича времето между момента на дразнене и началото на мускулното съкращение. Дължината му е много кратка - 0.002 – 0.004 секунди, тъй като бързината в една враждебна външна среда е от ключово значение за оцеляването на организма. По-силните дразнители водят до скъсяване на латентния период. След него започва съкращението на мускула. Следва фазата на разпускане. Накрая се наблюдават няколко малки единични трептения, резултат на мускулната еластичност. Периодът на съкращение на червените мускулни влакна е по-продължителен от този на белите. Мускулите на животните с постоянна телесна температура се съкращават по-бързо от тези на студенокръвните. Умореният мускул се съкращава много по-бавно от свежия. Наблюдава се и зависимост между силата на дразнене и степента на съкращение. Много слабите дразнители, които не довеждат до забележимо съкращение се наричат подпрагови. Най-малкият по сила дразнител, който предизвиква видимо съкращаване на мускула се нарича прагов. При естествени условия няма единични мускулни съкращения. При нормално функциониращ организъм се осъществяват продължителни съкращения, образувани от сливането на поредица единични съкращения. Те възникват в резултат на постъпващия залп от нервни импулси към мускула. Съкращението започва около 2 милисекунди след началото на деполяризацията на мембраната, преди реполяризацията да е завършила. Продължителността на съкращение при бързите мускулни влакна е около 7.5 милисекунди - те са свързани с фини, бързи и точни движения. При бавните мускулни влакна съкращението е от порядъка на 100 милисекунди - това са мускули, които извършват груби и продължителни движения, при които се развива голяма сила.
Изотоничното мускулно съкращение се изразява в това че мускулът се съкращава без съпротивление или при отсъствие на товар. В резултат на това напрежението в мускула не се променя, а се променя само дължината му. При изометричното съкращение на мускула се променя само напрежението, без да се променя дължината му. Практически се наблюдава, когато товарът е с тегло, което не е в състояние да се повдигне. Всички надпрагови дразнения водят само до промяна на напрежението без да се променя дължината му. Ако даден мускул се дразни с нарастваща честота на импулсите, в началото той ще успява да отговори на всяко дразнение с отделно съкращение. Когато се учестят обаче на 15 броя в секунда всяко следващо дразнене ще заварва мускула в не напълно разпуснато състояние преди да е завършил единичното съкращение. Фактически тетаничното съкращение се изразява в това, че мускулът се съкращава отново преди да се е разпуснал докрай. При тетаничното съкращение се наблюдава значителна загуба на вещества и енергия.
Приплъзването на тънките спрямо дебелите елементи на мускула води до неговото съкращение. Ширината на А-ивицата е постоянна, докато Z-линиите се приближават при съкращение на мускула и се раздалечават при разтягането му. Когато мусулът се скъсява, тънките филаменти от противоположните краища на саркомера се приближават, а при значително съкращение те се припокриват. Приплъзването при мускулното съкращение се осъществява като миозиновите глави се свързват здраво с актина, накланят се спрямо останалата част от миозиновата молекула, след което се отделят от актина. Този цикъл се повтаря многократно - при всяко едно мускулно съкращение. Структурата на главите на белтъка миозин 2 е била определена с помощта на кристалография с рентгенови лъчи. Всяка глава има актин-свързващо място и на 3.5 нанометра пред него - място за свръзване на АТФ. Последното е отворена цепка; при навлизане на молекула АТФ настъпва хидролиза и цепката се затваря. Това измества останалата част от главата, която се изправя - получава се тласък, който измества актина спрямо миозина. Всеки такъв тласък скъсява мускула с около 1 %. Схематично това изглежда така:
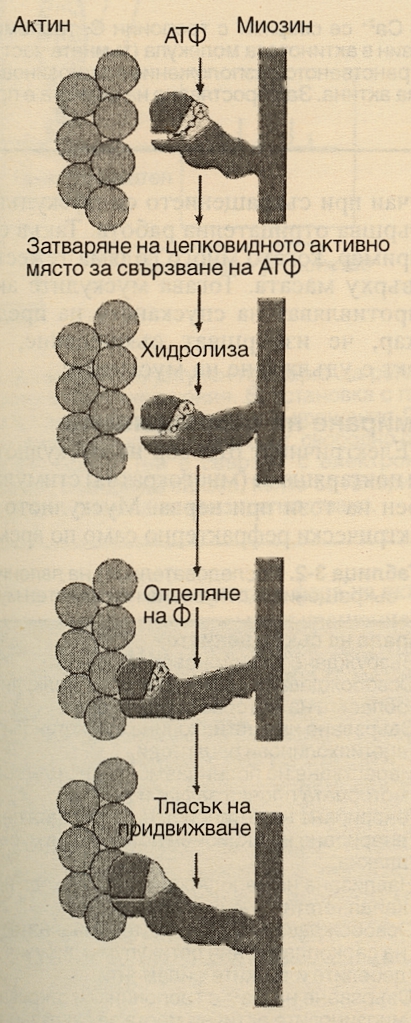
Всеки дебел филамент има приблизително 500 миозинови глави и при бързо съкращение се извършват по пет такива цикъла за секунда. Миозиновите глави имат ензимна активност - те извършват хидролизата на високоенергийния фосфат на АТФ. Процесът, при който деполяризацията на мускулното влакно инициира съкращение, се нарича куплиране на възбуждане и съкращение. Акционният потенциал се провежда до всички фибрили във влакното чрез Т-системата. Той отключва освобождаването на калциеви катиони от терминалните цистерни, страничните сакове на саркоплазмения ретикулум в близост до Т-системата. Калциевите катиони дават началото на съкращението - те се свързват с тропонин С. При отпуснат мускул тропонин I е здраво свързан с актина, а тропомиозинът покрива местата на свързване на миозиновите глави по актиновата молекула. Така комплексът тропонин - тропомиозин представлява релаксираща молекула, която инхибира взаимодействието между актина и миозина. Когато калциевите йони се свържат с тропонин С, връзаката на тропонин I с актина отслабва и това позволява на тропомиозина да се придвижва латерално. Графично това изглежда така:
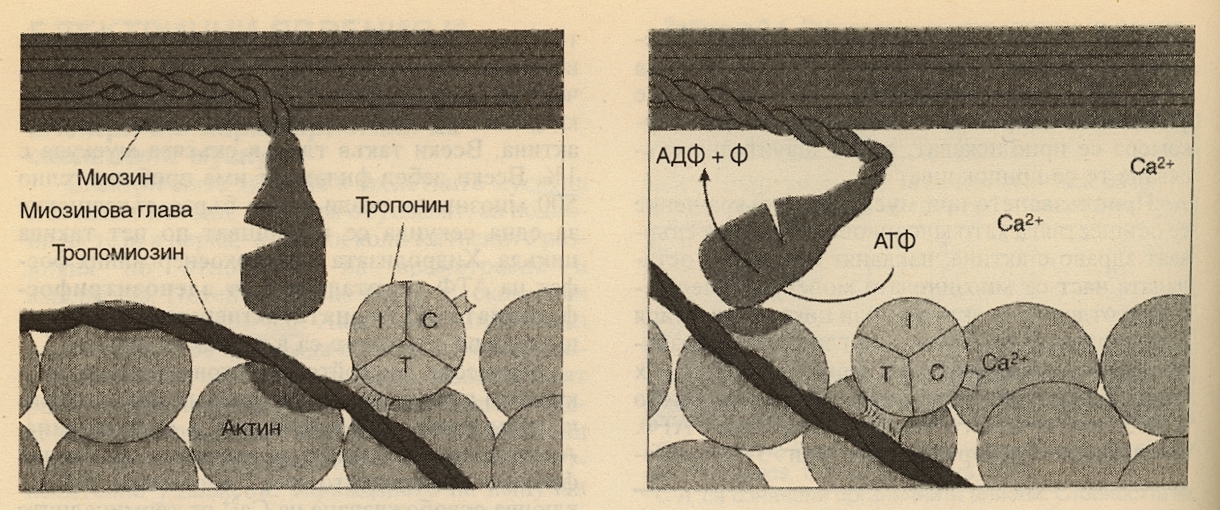
Това придвижване открива свързващите места за миозиновите глави. Тогава АТФ се разцепва и съкращението се осъществява - схемата вдясно. При свързването на всяка молекула тропонин с калций се откриват седем миозин - свързващи места.
Скоро след освобождаването на калциевите катиони те започват да се натрупват в лонгитудиналните части на ретикулума чрез активен транспорт. При този транспорт активно участие взема калциево - магнезиевата АТФ-аза. От там калцият дифундира в терминалните цистерни, където се складират до освобождаването си под действие на следващия акционен потенциал. При достатъчно понижаване на концентрацията на калция извън саркоплазмения ретикулум химичното взаимодействие между актина и миозина се прекратява и мускулът се отпуска. АТФ обезпечава енергийно както съкращението, така и отпускането. Ако транспортът на калций към ретикулума се блокира, настъпва трайно съкращение без възможност за релаксация мускулен спазъм. При мускулите - затварячи на долната челюст този спазъм носи названието тризмус - тъй като затварящите мускули са доста по-силни от отварящите, болният не може да си отвори устата.
Деполяризацията на мембраните на Т-тубулите активира саркоплазмения ретикулум чрез дихидропиридинови рецептори, които всъщност са волтаж - зависими калциеви каналчета в мембраните на Т-тубулите. При сърдечната мускулатура навлизането вътре в клетката на калциевите катиони през тези каналчета отключва освобождаването на същите йони от саркоплазмения ретикулум. При скелетните мускули обаче не е необходимо да навлизат йони от екстрацелуларната среда. Вместо това дехидропиридиновият рецептор изпълнява ролята на волтаж - зависим сензор и пусков механизъм, който отключва освобождаването на калций от близколежащите участъци на саркоплазмения ретикулум. Името на рецептора произлиза от възможността той да бъде блокиран от препарата дехирдопиридин. Рецепторът има четири хомоложни области, всяка от които пронизва Т-тубулната мембрана шест пъти. Калциевото каналче в саркоплазмения ретикулум, чието отваряне позволява изливането на калций, е наречено рианодинов рецептор, тъй като то се задържа в отворено положение от растителния алкалоид рианодин.
Мускулният тонус се изразява в способността на мускулите да поддържат, продължително и устойчиво състоянието си на леко съкращаване, при минимален разход на вещества и енергия. Чрез тоничното съкращаване на скелетните мускули се осигурява преодоляване на земното притегляне и поддържане на позата на главата и тялото. Тонусът се поддържа чрез непрекъснато изпращане на редки импулси от централната нервна система. Импулсите засягат несинхронно отделните мотоневрони, в резултат на което мускулът се съкращава по модела на слабо тетанично съкращение. Контрактурата се изразява в трайно съкращаване на отделни мускулни влакна и на мускула като цяло. Контрактурата бива вродена и придобита. Контрактурата от рефлекторен произход възниква като резултат от нарушенията на функцията на нервната система или в резултат на силна болка. Ако възникне в резултат на преумора се наблюдава увеличение на времетраенето на фазата на съкращението и особено на фазата на разпускането. Биохимично контрактурата на мускула се изразява в намаляване на веществата, участващи в плъзгането на миозиновите и актиновите нишки и връщането им в изходно положение.
Няколко часа след настъпване на смъртта мускулите се втвърдяват и това пречи на тяхната подвижност. Първоначално се вцепеняват мускулите на главата, следвани от тези на трупа и крайниците. Факторите, които влияят на контрактурата са същите и при следсмъртното вкочаняване, а именно образуването на тройна връзка между миозиновите и актиновите нишки като последица от изчезването на пластифициращото действие на АТФ.
Работата на мускулите се измерва с товара, който даден мускул повдига на определена височина. При увеличаване на натоварването в началото количеството на мускулната работа се увеличава. След достигане на максимума обаче тя започва значително да намалява и спада до нула. Началното увеличение на извършената работа се дължи на повишената възбудимост на мускула и на увеличението на степента на мускулното съкращение. Количеството на мускулна работа е правопропорционално на количеството на мускулните влакна и тяхната дължина. Върху количеството на извършената работа влияят разтягането на мускула под влияние на натоварването, количеството и разположението на мионевралния апарат. Уморените мускули извършват по-малко работа главно поради намаление височината на подема. Работата, която скелетните мускули извършват за да осигурят движението се нарича динамична. Напрежението на скелетните мускули, с което се осигурява преодоляването на земното притегляне и поддържане на тялото в пространството пък се нарича статично усилие. Мускулите могат да извършват отрицателна работа - което от физична гледна точка представлява твърдение без всякакъв смисъл. Такъв е случаят, при който много голяма тежест се спуска надолу под действие на гравитацията - тогава мускулите не могат да спрат тежестта, а само противодействат на нейното движение. При силовите тренировки това се нарича негативно повторение, но от физична гледна точка няма отрицателна работа - мускулното усилие забавя спускането на товара и противодейства на гравитацията. Има методи на тренировка, които включват само негативни повторения - което изисква някой асистент да повдигне тежестта, а трениращият само да забавя нейното спускане. В кръга на шегата по фитнес залите се говори че е необходимо някой шампион да помага на начинаещ - тъй като асистентът трябва да повдигне с бицепса си тежест, която е по-голяма от тежестта, която трениращият повдига от лежанка.
Функцията на мускулите е най-добре изразена при аеробни условия. Въпреки това за ограничено време те могат да функционират и при анаеробни - при липса на достатъчно кислород. След като работи така известно време и се постави отново в аеробни условия мускулът бързо поглъща кислород. С него се метаболизират обменните продукти образувани при мускулната активност при анаеробни условия. Способността на мускулите да работят при анаеробни условия се дължи на способността им да понасят кислородна недостатъчност. За една продължителна и ефективна мускулна активност е необходимо пълно възстановяване на кислородния дълг.
Коефициентът на полезно действие е съотношението на енергията употребена за работа на мускула към цялото количество енергия произведена от него. В най-добрия случай КПД се движи от 28 – 35 % - колкото е и КПД на двигателя с вътрешно горене. Върху него оказват влияние големината на товара и скоростта на мускулното съкращение. Той е най-голям при средно натоварване и не особено учестена работа. Мускулната сила се измерва с максималния товар, който даден мускул може да повдигне или с максималното напрежение, което той е в състояние да развие. Колкото по-голям е броят на влакната, толкова по-голяма е мускулната сила. Някои професионални състезатели по вдигане на тежести могат да повдигнат около 3 пъти по-голяма тежест от собственото си тегло над главата си - което изисква сериозна физическа подготовка и в повечето случаи - употребата на стероиди и други забранени стимуланти. При повдигане на щанга от лег (лежанка) за добро ниво на тренираност се приема способността човек да повдигне тежест около 1.5 пъти повече от собственото си тегло. Световният рекорд за повдигане на щанга от лег е 468 килограма - като състезателят, който го е поставил, е с тегло 160 килограма или отново 3 пъти по-малко от тежестта. Остава загадка дали е употребявал забранени стимуланти.
След период на интензивна активност мускулите губят способността си да се съкращават. Това състояние се нарича мускулна умора. Ако се прилагат продължителни стимули върху уморен мускул, то той достига до пълна липса на съкращение. Признаците на мускулна умора се изразяват в намаляване на амплитудата на съкращението и удължаване на времето му, което се простира върху трите фази. Много силната умора води до контрактура на мускула. Умората е нормален физиологичен процес, който води до прекратяване на работата. След време на почивка работоспособността на мускулите се възстановява. Съкращението на мускулните влакна няма рефрактерен период - повторната стимулация преди настъпването на отпускането води до допълнително активиране на съкратителния механизъм и отговор, който се прибавя към вече настъпилото съкращение. Това явление е известно като сумиране на съкращенията. При него развиваното напрежение е значително по-голямо от напрежението на единичното съкращение. При бързо повтаряща се стимулация активирането на контрактилните елементи настъпва всеки път преди те да се отпуснат и индивидуалните отговори се сливат в продължителен спазъм - тетанус или тетанично съкращение. Съществува пълен тетанус (когато между отделните съкращения не настъпва отпускане) и непълен тетанус (когато между сумиращите се стимули има частично отпускане). При пълен тетанус развиващото се напрежение е около 4 пъти по-голямо от това на единичното мускулно съкращение. Графично това изглежда така:

Съкращението е изометрично при постепенно нарастваща и намаляваща честота на стимулация. Точките бележат интервали от 0.2 секунди. Стимулиращата честота, при която настъпва сумиране на съкращенията, се определя от продължителността на единичното мускулно съкращение на мускулното влакно. Например при продължителност на единичното съкращение от 10 милисекунди честоти по-малки от 10 милисекунди предизвикват дискретни (отделни) отговори, които са разделени от пълно отпускане. При честота на импулса над 100 Hz (100 трептения в секунда) се стига до сумиране и тетаничен гърч.
Когато към скелетен мускул се приложи серия максимални стимули с честота, малко по-ниска от тетанизиращата, отначало се наблюдава покачване на напрежението, което се развива при всяко съкращение. След няколко съкращения се достига до постоянно за всяко съкращение напрежение. Това явление се нарича феномен на стълбата и се наблюдава и при сърдечния мускул. Смята се че се дължи на повишената способност на калциевите катиони да се свързват с тропонин С и не бива да се смесва със сумирането на съкращенията при тетанус.
Напрежението, което един мускул развива при изометрично съкрщаение, се нарича общо напрежение. Пасивното напрежение е упражнявано от нестимулиран мускул. Тези два показателя варират в зависимост от дължината на мускулното влакно. Дължината на мускула може да се променя при промяна в разстоянието между двете залавни места. Графично това изглежда по следния начин:
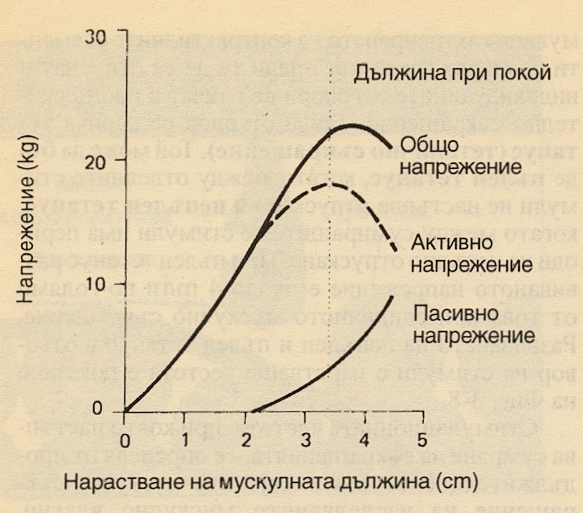
Зависимост между дължината и напрежението при човешкия мишничен мускул трицепс. Кривата на пасивно напрежение показва силата, която този мускул развива чисто пасивно - без да бъде стимулиран, единствено при свиване на лакътната става. Кривата на общо напрежение показва напрежението, което мускулът развива при изометрично съкращение в отговор на максимален стимул. Активното напрежение е резултатното между двете и се получава сумарно чрез интерпозицията на двете графики. Разликата между общото напрежение и пасивното напрежение се нарича активно напрежение. Дължината на мускула, при което активното напрежение е максимално, обикновено се нарича дължина на покой. В резултат на много експерименти е установено че дължината на повечето мускули в човешкия организъм при покой е дължина, при която те развиват максимално напрежение.
Наблюдаваната при скелетните мускули зависимост между дължината и напрежението се обяснява с механизма на приплъзване между филаментите по време на мускулното съкращение. Когато мускулното влакно се съкръщава изометрично, развиваното напрежение е пропорционално на броя на мостчетата между актиновите и миозиновите молекули. При разтегляне на мускула се намалява степента на припокриване между атина и миозина и по този начин намалява броят на напречните мостчета.
Скоростта на мускулното съкращение се променя обратнопропорционално на натоварването на мускула. При определен товар скоростта е максимална при дължина на покой и намалява ако мускулът е по-къс или по-дълъг от тази дължина.
Разтегливост е способността на мускулите да увеличават дължините си при натоварване и зависи от теглото на товара. Червените мускули са по-разтегливи от белите, а вретеновидните и паралелните са по-разтегливи от перестите. Еластичността е способността на мускулите да се възвръщат към първоначалното си състояние след деформация. По време на мускулните съкращения еластичността се увеличава, а разтегливостта му намалява. При умора разтегливостта се увеличава.
Мускулното съкращение се нуждае от енергия и всъщност мускулите са единствената тъкан в организма, която превръща химичната енергия в механична работа. Непосредственият източник на тази енергия са енергитично богатите органични фосфорни съединения в мускула, а решаващият източник е междинната обмяна на въглехидрати и липиди. Аденозинтрифосфатът се синтезира от АДФ чрез присъединяване на фосфатна група. Част от енергията за тази ендотермична реакция се доставя от разцепването на глюкоза до въглероден диоксид и вода. Друго съединение, което може да обезпечи енергийно съкращението за кратко време, е креатинфосфатът. Той се хидролизира до креатин и фосфатни групи с освобождаване на значително количество енергия. При покой част от АТФ прехвърлят в митохондриите фосфата си към креатина и така се изгражда запас от креатинфосфат. По време на съкращение креатинфосфатът се хидролизира в местата за свързване на миозиновите глави с актина и се образува АТФ от АДФ. Това позволява съкращението да продължи.
При покой и при леко физическо натоварване мускулите използват липиди под формата на свободни мастни киселини. Когато интензивността на физическите упражнения нараства, липидите не могат да доставят енергия достатъчно бързо и става необходимо да се използват въглехидрати. Така по време на физически упражнения по-голямата част от енергията, необходима за ресинтезата на фосфокреатина и АТФ, идва от разграждането на глюкоза до въглероден диоксид и вода. Глюкозата от кръвообръщението навлиза в клетките, където чрез серия от метаболитни процеси се разгражда до пируват. Друг източник на вътреклетъчна глюкоза, респективно на пирувт, е гликогенът - въглехидратен полимер, който се открива в голямо количество в черния дроб и скелетните мускули. В присъствие на достатъчно кислород пируватът навлиза в цикъла на лимонената киселина, при което се метаболизира с помощта на така наречените дихателни ензимни пътища до въглероден диоксид и вода. Този процес се нарича аеробна гликолиза. При метаболизма на глюкозата до въглероден диоксид и вода се освобождава значително количество енергия - тя се складира под формата на съединението АТФ, което се синтезира от АДФ. Ако снабдяването с кислород е недостатъчно, пируватът, образуван от глюкозата, не навлиза в цикъла на трикарбоновите киселини, а се редуцира до лактат. Този процес на анаеробна гликолиза в свързан с чистата продукция на много по-малко количество енергитично - богати фосфатни връзки, но не се нуждае от кислород.
По време на мускулната работа кръвоносните съдове в мускулната тъкан се разширяват и притокът на кръв към мускула се увеличава - което води до увеличаване на доставката на кислород. До определен момент нарастването на кислородната консумация е пропорционално на необходимата енергия и всички енергитични потребности се задоволяват аеробно. При по-голямо мускулно усилие обаче аеробната ресинтеза на енергийните запаси не може да се изравни с тяхното използване. При тези условия креатинфосфатът все още се използва за ресинтеза на АТФ. Известно количество АТФ се получава при анаеробното разграждане на гюкозата до лактат (млечна киселина). Използването на анаеробните пътища обаче е самоограничаващ се процес, тъй като независимо от бързата дифузия на лактата до в кръвообръщението в мускулите се натрупват достатъчно количества. Те могат да надхвърлят капацитета на тъканните буфери и да доведат до понижаването на рН - все пак млечната киселина има кисела реакция. Този процес води до инактивация на ензимите; за кратко време обаче този процес позволява да се извърши много по-голямо количество работа, отколкото тя би била възможна при липса на анаеробни пътища за разграждане. Например при бягане на къси разстояния (100 метра за 10 секунди) около 85 % от необходимата енергия се доставя анаеробно. При пробягване на 3 километра за 10 минути само 20 % от енергията се доставя анаеробно, а при бягане на маратонски и полумаратонски дистанции (за време от порядъка на 60 и повече минути) само 5 % от необходимата енергия се доставя по анаеробен път. При силовите тренировки натоварването е основно анаеробно - рядко някое упражнение включва повторения за повече от 10 секунди. Това налага включването и на упражнения, включващи аеробен режим - колоездене, бягане (на пътечка или крос), изкачване на планини и т.н. На фитнес жаргон тези упражнения се наричат кардио - серии, тъй като се смята че стимулират сърдечната дейност. По време на такива упражнения се стига до ускорено разграждане на липидите.
След приключване на натоварването кислородната консумация остава по-висока от нормалното. Така се отстранява излишъкът от лактат, попълват се запасите от АТФ и се насища миоглобинът с кислород. Количеството на кислородната свръхконсумация е пропорционално на степента, в която енергитичните потребности по време на физическата работа надхвърлят капацитета на механизмите на аеробен синтез на енергийни резурси - кислороден дълг. Експериментално кислородният дълг се измерва чрез определяне на кислородната консумация след натоварване до достигане на постоянната (изходна) стойност и изваждане на изходната от тоталната консумация. Количеството на кислородния дълг може да надхвърли 6 пъти изходната кислородна консумация, което показва че човекът е способен да извърши шест пъти по-голямо физическо натоварване, отколкото би било възможно без механизма на кислородния дълг. Максималната стойност на този дълг може да се постигне бързо или бавно; екстремни натоварвания са възможни само за кратко време, докато по-лека физическа работа може да се извършва и за по-дълги периоди.
Тренираните атлети са в състояние да увеличават кислородната консумация на мускулите си в по-голяма степен от нетренираните хора. Като резултат те са в състояние да извършват по-големи физически натоварвания без да изчерпват гликогеновите си резерви и без да увеличават продукцията на млечна киселина. Именно лактатът води до така наречената мускулна треска - болки и леки изтръпвания в мускулите след натоварване, особено около 24 часа след това. При една и съща мускулна работа тренираният човек натрупва по-малък кислороден дълг от нетренирания. Няколко дни преди състезания активните спортисти започват да консумират големи количества въглехдрати, което натрупва енергийни резерви в мускулите; при културисти това води дори до известно увеличаване на мускулния обем, но не бива да се прекалява - въглехидратите бързо се метаболизират до мазнини, подкожната мастна тъкан се увеличава като количество и мускулният релеф се губи.
При пълното изчерпване на запасите от АТФ и креатинфосфат в мускулите се развива състояние на крайна ригидност - ригор. Когато това състояние настъпи след смъртта, то носи названието rigor mortis - трупно вкочаняване, и може да си използва от криминалистите за установяване на времето, преди което е настъпила смъртта. В състояние на ригор почти всички миозинови глави са свързани с актина устойчиво.
Според законите на термодинамиката енергията, доставяна на мускула, трябва да бъде равна на енергията, изразходвана от него. Енергийните разходи включват работата, извършвана от мускула, макроергичните фосфатни връзки, образувани с цел по-ксъно използване, както и произведената от мускула топлина. Коефициентът на полезно действие на скелетния мускул (извръшена работа / обща изразходвана енергия) е около 50 % при изотонично и 0 % при изометрично съкращение. От физична гледна точка обаче в много случаи изометричното съкращение изисква значителен разход на енергия с цел поддържането на статично положение на тялото, така че последното твърдение просто не отговаря на истината.

Схема на упражнението планк - може би най-често използваното статично натоварване във фитнеса. Упражнението натоварва делтоидните мускули, коремната преса, широкият бедрен мускул и предната част на подбедрицата. В общи линии статичните упражнения не са особено приятни за изпълнение - водят до тъпа, неприятна болка, изтръпване и много тениращи бързо се отказват. Това е довело до появата на голям брой мемета и всякакъв хумор, който само един активно трениращ може да осмисли на практика:

Натрупването на енергийни резерви под формата на фосфатни връзки се отразява слабо на енергоразхода - което означава че продукцията на топлина от мускилите е значителна. Произведената от мускула топлина може да бъде измерена чрез подходяща термодвойка.
Топлината на покой е резултат от основните метаболитни процеси. По време на съкращение се произвежда начална топлина. Тя се състои от активационна топлина и топлина на скъсяване - нейното количество е пропорционално на величината на скъсяване на мускула. След мускулното съкращение производството на топлина продължава около 30 минути. Това представлява топлината на възстановяване, освободена от метаболитните процеси, които връщат мускула в състоянието му на покой преди съкращението. Топлината на възстановяване на мускула е приблизително равна на неговата първоначална топлина, т.е. топлината на възстановяване е приблизително равна на произведената по време на съкращението топлина. Поради това всеки, който започне силови тренировки, забелязва как през първите няколко месеца устойчивостта на организма към по-ниски температури рязко се увеличава. Ако мускул, който се е съкратил изотонично, се върне към изходната си дължина, освен топлината на съкращение се генерира и топлина на отпускане. Последната е израз на външната работа, която трябва да се извърши, за да се върне мускулът в изходното си състояние.
При здрави животни и хора скелетните мускули се съкращават единствено при стимулация от страна на инервиращите ги моторни неврони. При деструкция на тези нерви настъпва афункционална атрофия - която може да се предотврати със съответната рехабилитация. В тези случаи първоначално се наблюдава абнормна възбудимост на мускулите и покачване на чувствителността им спрямо циркулиращия ацетилхолин (денервационна хиперсензитивност). Появяват се фини неритмични потрепвания на отделни влакна (фибрилации). Тези явления изчезват при процеса на регенерация на двигателните нерви. Такива съкращения обикновено не са ясно видими и не бива да се смесват с фасцикулациите - последните са видими потрепвания на мускулните влакна, дължащи се на патологични разряди на спиналните мотоневрони. Постепенно фибрилациите изчезват и настъпва атрофия - именно този процес може да се предотврати с помощта на различни рехабилитационни процедури, тъй като при по-възрастни пациенти атрофията е необратима и трайно инвалидизира организма.
Всеки аксон от спиналните мотоневрони се разклонява и инервира няколко мускулни влакна. Поради това най-малката част от мускула, която може да се съкращава самостоятелно, не е единично мускулно влакно, а снопче от няколко влакна, инервирани от един неврон. Това снопче носи названието двигателна единица. Броят на мускулните влакна във всяка една двигателна единица е различен. При мускули, които извършват фини движения и се изисква отлична координация, една двигателна единица съдържа 3 - 6 мускулни влакна. Такива са мускулите на ръката, очедвигателните мускули, мускулите на езика и ларинкса. От друга страна, при някои големи по размер мускули двигателните единици съдържат много голям брой мускулни влакна - 120 - 160, дори 200 броя. Такива са широките гръбни мускули и четириглавият бедрен мускул при човека.
Всеки спинален мотоневрон инервира само един тип мускулни влакна, така че всички мускулни влакна в една двигателна единица са от един и същи тип. Следователно двигателните единици се подразделят на бързи и бавни, точно както и мускулните влакна. Най-общо бавните мускулни единици са инервирани от малки, бавнопровеждащи двигателни неврони, докато големите - от големи, бързопровеждащи неврони - принцип на размера. В големите мускули на крайниците най-често използваните единици, които се включват и първи, са малките, бавни единици, които са устойчиви на умора. Бързите единици, които са лесно уморяеми, се включват главно в движенията, изискващи повече сила.
Разликите между отделните типове мускулни влакна (съответно единици) не са неизменно присъщи. Ако нервът към бавен мускул се пререже и се замени с нерв към бърз мускул, нервът регенерира и инервира бавния мускул. Мускулът тогава се превръща в бърз, при което настъпват и съответните промени в изоформите на мускулните белтъци и активността на аденозинтрифосфатазата. Съществуват биохимични проучвания, според които тези промени се дължат на трофични субстанции, които по хода на нерва достигат до мускула. Като краен резултат се променя характерът на мускулната активност. В опити със стимулация е показано че промени в характеристиката на използваната електрична стимулация могат да предизвикат промени в експресията на МТВ-гените, респективно в МТВ-изоформите.
Денервацията на скелетния мускул води до атрофия и вяла парализа с появата на фибрилации. Тези ефекти са класически последици от увреждането на долните мотоневрони. Мускулът освен това става хиперсензитивен към ацетилхолин. Всички тези промени могат да се регистрират с електромиография - измерване и записване на електричната активност на мускула с катодно - лъчев осцилоскоп. Регистрацията може да се извърши чрез поставяне на малки дисковидни метални електроди на кожата над изследвания мускул, а също и с помощта на отвеждащи игловидни субкутанни електроди. Полученият по този начин запис се нарича електромиограма и понякога се извършва с диагностична цел подобно на електрокардиограмата. С тънки игловидни електроди може да се регистрира електричната активност дори на единични двигателни единици. Електромиографията на човешки двуглав и триглав мишничен мускул (съответно бицепс и трицепс) изглежда така:
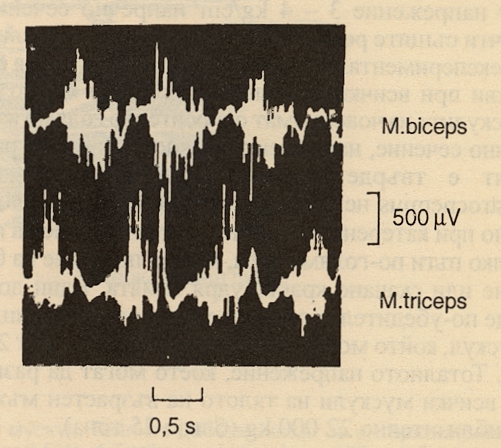
Движенията, при които се получава подобна активност, са флексия и екстензия в лакътната става. Важно е че при липса на патологични процеси мускулите на човека имат минимална или нулева спонтанна активност. При минимална волева активност се активират няколко двигателни единици, а при нарастващо волево усилие се активират все повече и повече - този процес се нарича рекрутиране на двигателните единици. Поради това градирането на мускулния отговор е отчасти функция на броя на активираните двигателни единици. Допълнителна роля играе честотата на разрядите на единичните нервни влакна. При описаното по-горе тетанично съкращение развиваното мускулно напрежение е по-голямо от нормалното волево съкращение - пациенти в гърчови състояния развиват огромна сила и са в състояние да разскъсат дрехи, колани и дори белезници - нещо, което е непостижимо при нормални условия за същия индивид. Значение има и дължината на мускула. Различните двигателни единици се възбуждат асинхронно с разместване на фазите - което води до плавно съкращение, а не такова на тласъци.
Човешките мускули могат да създават напрежение от порядъка на 3 - 4 килограма на квадратен сантиметър напречно сечение. Почти същите резултати са постигнати при всички бозайници. Тъй като много от мускулите при човека имат голямо напречно сечение, те могат да развиват и голяма сила (напрежение). Така например musculus gastrocnemius не само че поема тежестта на цялото тяло при катерене, но оказва и голямо напрежение върху земната повърхност когато кракът удря земята. Големият седалищен мускул пък е възможно да повдигне тежест от около 1200 килограма - стига тежестта да бъде прикрепена към мускула по подходящ начин от гледна точка на биомеханиката. Тоталното напрежение, което могат да развият мускулите на възрастен мъж, се изчислява на приблизително 22 тона.
Движенията на човешкото тяло в триизмерното пространство се извършват по такъв начин, че се използват максималните предимства от всички вече описани биомеханични принципи. Например мускулите са прикрепени по такъв начин, че много от тях в началния момент на съкращението са с близка дължина до дължината на покой - не са преразтегнати. При мускули, които преминават през повече от една става, движението на първата става може да компенсира движението на втората по такъв начин че сумарното съкращение на мускула да бъде всъщност минимално - така се постига максимална ефективност с минимална сила и се спестява енергия. Някои от бедрените мускули преминават над тазобедрената става и под тибията и фибулата, при което съкращението им предизиква флексия на подбедрицата спрямо бедрото. Ако в същото време бедрото е фиксирано за таза, удължаването на тези мускули в тазобедрената става може да компенсира скъсяването им при колянната става. По време на различни форми на активност тялото се движи така че извлича предимствата от тази биофизична закономерност. Фактори като момент и баланс се интегрират към телесните движения така че правят възможно максималното движение с минимално мускулно усилие. В крайна сметка напрежението, приложено върху сухожилията и костите, рядко достига до 50 % от увреждащата ги стойност, което ги предпазва от наранявания.
При ходене всеки крайник преминава ритмично през поддържаща фаза, когато кракът е на земната повърхност, и през висяща фаза, когато кракът е отделен от нея. Поддържащите фази на двата крака се припокриват, така че при всеки цикъл има два периода на двойно поддържане. В началото на всяка стъпка има залп на активността на флексорите, след което кракът се повдига напред чрез много малко допълнително съкращение. Следователно мускулите са активни само през част от времето на всяка стъпка, поради което и продължителното ходене е свързано с относително малка умора.
Израстналият млад човек при ходене с нормално темпо изминава около 80 метра за една минута или в рамките на 4 - 6 километра за един час. Генерираната мощност е от порядъка на 150 - 175 вата за всяка една стъпка. Това е най-удобното темпо за ходене - по-бързият или по-бавният ход изморяват повече и изискват повече енергия. Въпреки че ходенете е сложна двигателна активност, то е до голяма степен автоматизирано. То се организира от предварително програмирани двигателни пътища по гръбначния мозък и се активира от командни неврони - подобни са физиологичните механизми при човека и различни животни, като при безгръбначните в най-общия случай координацията е опростена.
Мускулната атрофия на Дюшен и по-леко протичащата мускулна дистрофия на Бекер са вродени заболявания, свързани с Х-хромозомата. Те се характеризират с прогресиращо мускулно отслабване. Мускулната дистрофия на Дюшен се среща повече при мъжкия пол, при едно от около 3000 момчета, като около тридесетата година от живота на човека обикновено води до летален изход. Изолиран е ген, който при болните индивиди е дефектен и води до синтезата на белтъка дистрофин. Това е голям протеин с молекулно тегло около 427 000 единици и има формата на издължена пръчица. Той може да свързва вътрешния скелет на мускулните клетки с други клетки и по този начин позволява координирането на мускулното съкращение. Прогресивната загубана мускулната функция се дължи предимно на загубата на тази координация. При мускулната дистрофия на Дюшен белтъчната молекула изобщо липсва, докато при болестта на Бехер тя е с неправилна форма и ниски нива на концентрация в мускулната тъкан. Генът, кодиращ дистрофина, е един от най-големите човешки гени. Експресира се и в сърдечния, и в гладките мускули, а с алтернативния промотор и първи екзон - и в мозъка.
При различните форми на мускулните миотонии волевото съкращение се последва от удължена фаза на отпускане. Миотониите се дължат на дефектни гени в хромозоми 7, 17 и 19, които предизвикват отклонения от нормата на натриевите или калциевите каналчета.
При ембрионалното развитие на мускулите основна роля играе транскрипционният фактор миогенин. Той индуцира превръщането на фибробластите в мускулни клетки. При раждане на мишки, хомозиготни по отношение на мутантния ген, кодиращ миогенин, те умират поради липсата на мускули - включително и тези, необходими за дишането.
Всички мускули се образуват от параксиалната мезодерма на ембриона. Те се разделят по дължината му на сомити, съответстващи на сегментирането на тялото, което е най-лесно забележимо при гръбначния стълб. Всеки сомит има три части – склеротом, от който се образува прешленът, дерматом, от който се образува кожата и миотом, от който се образуват мускулите. Миотомът от своя страна се разделя на две части, епимер и хипомер, от които се образуват съответно епаксиални и хипаксиални мускули. При хората епаксиални мускули са само дългият гръбен мускул и малките междупрешленни мускули. По време на развитието на зародиша миобластите или остават в сомита, за да образуват мускулите около гръбначния стълб, или мигрират навън в тялото, за да образуват всички останали мускули. Миграцията на миобластите е предшествана от образуването на структура от съединителна тъкан. Миобластите се придвижват към местата си, следвайки химични сигнали, като при достигането на съответното анатомично място започва процес на срастване в удължени скелетни мускулни клетки.
Лейомиогенезата (на латински: leiomyogenesis) се осъществява от два източника. Основната маса на гладката мускулатура изграждаща вътрешните органи произлиза от мезенхима, получен от висцералната мезодерма. Миоепителните гладкомускулни клетки възникват от ектодермата. Залагането им във вътрешните органи става на определени места като струпване на мезенхимни клетки (лейомиобласти), които постепенно се удължават, а в цитоплазмата им се появяват протомиофибрили. Първоначално те се свързват мрежовидно, но по-късно се прилепват помежду си плътно.
Рабдомиогенезата (на латински: rhabdomyogenesis) представлява образуване на мускулите по време на ембрионалното развитие и е важен етап от формирането на човешкия организъм. Мускулите на трупа се образуват от мезенхима, получен от миотомите. Мускулната тъкан на крайниците се образува от миотомите в областта на шията и пояса. Скелетно-мускулните влакна представляват симпласти, които произлизат от мезенхима. Чрез деление, разрастване и диференциране се превръщат в миобласти. Това са вретеновидни клетки, в чиято цитоплазма се появяват първични миофибрили. Те притежават контрактилна способност още в ранна ембрионална фаза. Миобластите притежават по едно ядро, което впоследствие се дели многократно. Увеличава се и количеството на цитоплазмата, натрупва се гликоген и се увеличава броят на миофибрилите без да се дели тялото на мускулното вретено. Сърдечно-мускулните клетки и влакна произлизат от мезенхимните клетки на миоепикардиалната плоча. Те постепенно загубват своите израстъци, удължават се нишковидно и се превръщат в цилиндрични образувания. Те се допират със своите краища и се сливат в дълги цилиндрични образувания, наречени сърдечно-мускулни влакна. Сърдечно-мускулните влакна не се натрупват в снопчета, а се свързват мрежовидно.
Почти няма област на хирургията, в която да не се работи в пределите на мускулната тъкан. Дори и оралната хирургия много често изисква обработка на мускулната тъкан - основно напречнонабраздена. Поради това всеки хирург трябва да познава някои чисто клинични особености на мускулите, важни за ежедневната практика.
Анатомия на мускулите
Мускулно съкращение
Болести на мускулите
Ембрионално развитие на мускулите
Мускулите и хирургията